Traducción del documento original Infant Calming Responses en exclusiva para Mimos y Teta. Prohibida su copia y distribución sin autorización expresa de Nohemí Hervada
Resumen
Contexto: El vínculo madre-bebé es la relación social más temprana y crítica de las crías de mamíferos. Para promover este vínculo, los recién nacidos poseen comportamientos innatos para buscar la proximidad materna y protestar contra la separación comunicándose con la madre verbalmente y a través del movimiento corporal. Sin embargo, los mecanismos fisiológicos que regulan estos comportamientos infantiles continúan estando ampliamente indefinidos.
Resultados: Aquí mostramos un innovador conjunto de respuestas cooperativas infantiles durante el embarazo. Los bebés menores de 6 meses de edad porteados por una madre caminando, cesaron inmediatamente el movimiento voluntario y el llanto, y mostraron un descenso rápido del ritmo cardíaco, comparado con aquellos sostenidos por una madre sentada. Es más, identificamos respuestas impresionantemente similares en crías de ratón definidas por la inmovilidad y las vocalizaciones ultrasónicas y ritmo cardíaco disminuidos. Empleando intervenciones farmacológicas y genéticas en las crías de ratón, identificamos los sistemas neurales que regulan la respuesta calmante. Se requieren entradas de señalización somatosensorial y proprioceptiva para la inducción, y las funciones parasimpáticas y cerebelosas median los gastos cardíacos y motores respectivamente. La pérdida de la respuesta de calma obstaculizó el rescate materno de las crías, sugiriendo una importancia funcional de las respuestas de calma identificadas.
Conclusiones: Nuestro estudio ha demostrado por primera vez que la respuesta de calma infantil al porteo materno es un conjunto coordinado de regulaciones centrales, motores y cardíacas y un componente conservado de interacciones madre-bebé. Nuestros descubrimientos proporcionan evidencia para, y tienen el potencial de, impactar la teoría y práctica actuales sobre la crianza, ya que el llanto no mitigado es el gran factor de riesgo de abuso infantil.
Introducción
Los bebés mamíferas requieren de un cuidado parental continuo para la supervivencia y el desarrollo psicosocial. Para lograr esto, los bebés poseen el impulso innato para buscar la proximidad materna y protestar por la separación, comunicándose con la madre vocalmente y a través del movimiento corporal. Estas contribuciones activas por parte de los bebés son tan esenciales como los comportamientos paternos que proporcionan los cuidados para la creación del vínculo madre-bebé. Aún así, a pesar su crítica importancia, los mecanismos fisiológicos y sensomotores que regulan estos comportamientos en los bebés son comprendidos de manera vaga, especialmente las respuestas positivas que median en los bebés, como la calma y la relajación en respuesta al cuidado maternal.
Los mamíferos nidícolas neonatos tienen una habilidad ambulatoria limitada y requieren porteo materno para su transporte. En los humanos, portear un bebé en brazos, en portabebés o en un carricoche mientras se anda, se realiza a menudo como una medida reconfortante. Sin embargo, el efecto calmante del porteo infantil ha sido controvertido en los estudios previos. (4-7), todos los cuales midieron la cantidad total de llanto y porteo durante una hora o más, de acuerdo a los diarios de los padres, y no a la observación directa, e intentaron correlacionar estas variables. Es más, en esos informes no se hizo distinción entre el porteo móvil y el simple soporte sin movimiento. En este estudio, nos centramos en los graves efectos a tiempo real del porteo materno, con monitores de audio-video y electrocardiogramas, que nos permitieron grabar los comportamientos de los bebés y las respuestas fisiológicas en una escala de tiempo por debajo de un segundo. Usando estos métodos, investigamos las respuestas infantiles al porteo materno y los mecanismos sensomotores tanto en los bebés humanos como en las crías de ratón, para dilucidar el comportamiento cooperativo infantil durante el porteo materno en los mamíferos.
Resultados
Las respuestas de calma al porteo materno en los bebés humanos
El comportamiento, vocalización, y electrocardiograma (ECG) de bebés humanos fueron monitorizados durante tareas de comportamiento que consistían en estar tumbados en una cuna, (CUNA), sostenidos por la madre sentada en una silla (sostenido), o sostenido por la madre mientras caminaba continuamente (porteo) (Figura 1ª). En experimentos iniciales pusimos a prueba varias duraciones y combinaciones de las tres condiciones (Figuras S1A y S1 disponibles online), y encontramos que todas las medidas del bebé (llanto, movimiento corporal y ritmo cardíaco) incrementaron generalmente durante la condición CUNA mientras que disminuyeron durante el porteo. La condición sostenido tuvo efectos intermedios entre las de CUNA y porteo. Durante las repeticiones sostenido-porteo (Figura 1 y Clip S1), el intervalo entre latidos (lo contrario del ritmo cardíaco) aumentó rápidamente al principio del porteo y volvió al nivel previo tras el principio del siguiente sostenido. Por lo tanto, en este estudio nos centramos en el periodo de transición sostenido-porteo.
12 bebés sanos, de edad entre 1 y 6 meses (media= 3.08 +- 0.51 meses, seis hembras y seis varones) fueron reclutados para los experimentos principales. Diez minutos después de aplicar electrodos holter, se pidió a las madres que realizaran una de las tres condiciones (CUNA, sostenido o porteo), secuencialmente, durante 30 segundos cada una, en orden aleatorio indicado por el experimentador. Subsecuentemente la información de los bebés despiertos durante el período de tiempo del sostenido seguido del porteo fue recopilada y analizada. Los bebés lloraban aproximadamente durante la mitad de la condición de sostenido inicial, (Figura 1D). Cuando la madre se levantó y comenzó a caminar, los bebés detuvieron los movimientos voluntarios significativamente. (t, prueba de los estudiantes), y el llanto (t) (Figuras 1C y 1D y Tabla S1). Es más, encontramos que el intervalo entre latidos de los bebés se elevaba inmediatamente después del principio del porteo (media +- SEM) = 7.39% … constante temporal 3.16s, corresponde a ocho cuentas de latidos de corazón…) (Figura 1E).
El llanto infantil aumenta seriamente la actividad física y altera significativamente el patrón respiratorio. Por lo tanto, el aumento del intervalo entre latidos podría ser derivado del cese del llanto. Para abordar esta cuestión, clasificamos cada episodio de transición sostenido a porteo en cuatro grupos: (1) los bebés no lloraron durante todo el periodo (n=25), (2) los bebés lloraron más del 50% del tiempo durante el sostenido pero no durante el porteo (n=13), (3) los bebés lloraron más del 50% del tiempo durante el sostenido y el porteo (n=6), y (4) otros (excluidos de este análisis; n=8). Los cambios en los intervalos entre latidos fueron analizados en estos grupos (Figuras 1F-1H). El mayor cambio se observó en el grupo de bebés que lloraron solo durante el sostenido (t…; Figura 1F). Sin embargo, el rápido aumento en el intervalo entre latidos debido al porteo se observó también en bebés que no lloraron durante el sostenido (aumento (media….), (Figura 1G), indicando que al menos alguna porción del efecto cardíaco era independiente del cese del llanto. Para el grupo 3, los bebés lloraron 96.7%…) durante el sostenido y 65.8… durante el porteo, y los intervalos entre latidos aumentaron significativamente tras el principio del porteo (t… ) (Figura 1H). Un modesto descenso de la frecuencia fundamental del llanto F(0) se observó tras el inicio del porteo en el último grupo de bebés (t…) (Figura 1). Un análisis aparte mostró que la elevación del intervalo entre latidos debido al porteo no se observó en bebés que dormían (Figura 1J y Tabla S1), lo que sugiere un efecto techo o la implicación de la función cognitiva o conciencia del bebé. Tanto en los bebés despiertos como en los dormidos, se observó la reducción transitoria del intervalo entre latidos (Figura 1G, de –9 a 0 cuenta cardíaca; Figura 1J, de -4 a 0 cuenta cardíaca). El tiempo inicial de esta reducción del intervalo entre latidos corresponde aproximadamente al inicio de los movimientos maternos de preparación para levantarse, tales como reposicionar al bebé. Por lo tanto, estas bajadas transitorias del intervalo entre latidos se podrían explicar como un reflejo defensivo del bebé (aceleración del ritmo cardíaco causado por el bebé poniéndose en alerta debido a una estimulación repentina e intensa), combinado con un baroreflejo causado por el levantamiento materno similar a la prueba de inclinación de la cabeza boca arriba. Sin embargo, la elevación mantenida de los intervalos entre latidos debido al porteo en los bebés despiertos no pudo ser explicada por ningún reflejo cardíaco vagal conocido, incluido el reflejo orientativo (breve periodo de deceleración del ritmo cardíaco por estímulos sensoriales leves) (8), lo que sugiere que el porteo evoca una reducción mantenida del ritmo cardíaco en conjunto con los rápidos cambios de comportamiento en los bebés humanos a través de un mecanismo innovador.
Análisis de variabilidad del ritmo cardíaco revelaron que el índice de actividad parasimpática rMSSD (raíz cuadrada de la media de la suma de los cuadrados de las diferencias entre los intervalos entre latidos adyacentes durante la duración del análisis) (9), era significativamente mayor durante el porteo que durante el sostenido (t…) (figura 1K). Estos datos sugieren que los bebés estaban más relajados durante el porteo que durante el sostenido, no solo en su comportamiento, sino también fisiológicamente.

Respuestas de calma al porteo en crías de ratón
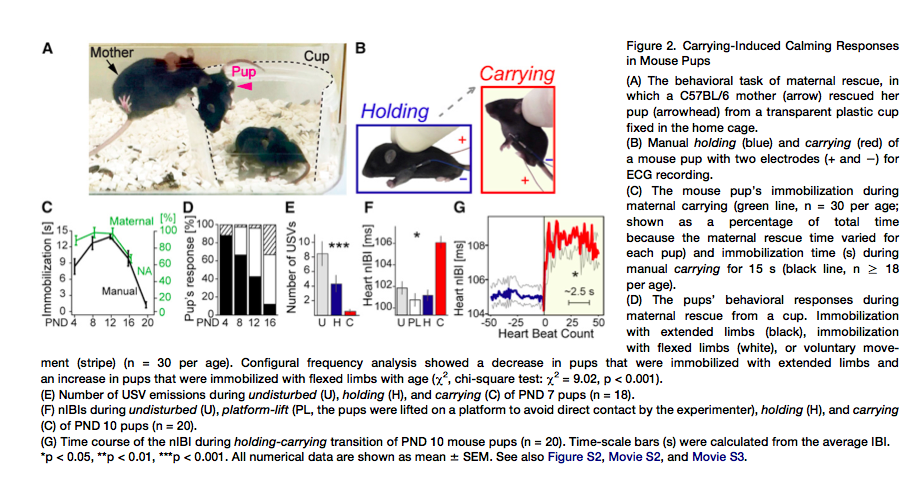
Nuestros descubrimientos con bebés humanos son evocativos de la respuesta al transporte materno oral observado en jóvenes mamíferos nidícolas, como los gatos y las ardillas. Los jóvenes porteados adoptaban a menudo una característica postura compacta con sus patas traseras encogidas (10,11). Sin embargo, no se han llevado a cabo experimentos para medir directamente el comportamiento calmante inducido por el porteo y los cambios fisiológicos concomitantes en los jóvenes porteados. Propusimos la hipótesis de que, similarmente a los bebés humanos, las crías de ratón también tendrán respuestas en el comportamiento y fisiológicas que acompañan el cambio en la postura en respuesta al porteo materno. Para probar la hipótesis, diseñamos una tarea de comportamiento naturalista en el que la madre ratón rescataba crías metidas en una copa (Figura 2A y clip S2). Cada día durante los días postnatales (PNDs) 4-16, se sacaron tres crías del nido y fueron colocadas en una copa para inducir el rescate materno. Todas las crías fueron recuperadas en menos de 15 minutos. Las respuestas de las crías a la recuperación materna y al rescate materno fueron medidas a través de un video análisis fotograma a fotograma. Encontramos que durante el porteo materno, las crías mantuvieron una postura inmóvil y compacta (Figuras 2C y 2D) durante la mayor parte de la duración de la tarea. La respuesta calmante ya no era evidente en el PND20, cuando las crías se estaban destetando y eran capaces de escapar de la copa por sí mismas (datos no mostrados). Respuestas similares podrían ser inducidas también por el porteo manual de un experimentador, sosteniendo la pequeña cantidad de piel de la nuca, imitando el agarre oral materno (Figuras 2B y 2C y Clip S3).
A continuación examinamos si la respuesta calmante de las crías portadas era similar a aquella de los bebés humanos. Las crías roedoras emiten vocalizaciones ultrasónicas (USVs) a 40-80 kHz cuando son separadas de sus madres y el resto de la camada (12). Las madres ratón se dirigieron a grabaciones digitales de USVs de crías, pero no a ningún otro ultrasonido sintetizado (13), lo que sugiere una función de las USVs comparable al llanto en los bebés humanos (14). Encontramos que el porteo redujo rápidamente la emisión de USV de las crías comparado con el sostenido o condiciones sin estorbos. (F, Fisher´s ANOVA…) (Figura 2E). Es más, similar a los bebés humanos, el porteo tuvo un efecto significativo en el aumento del intervalo entre latidos de las crías (aumento (media…., constante temporal = 0.55 s; correspondiente a 5 cuentas de latidos de corazón; t….) (Figuras 2F, 2G y S2). El aumento de intervalo entre latidos no ocurrió cuando las crías eran sostenidas del mismo modo, pero sin ser levantadas, o cuando las crías eran levantadas junto con una bandeja inferior de plástico (plataforma) (Figura 2F). Por lo tanto, en las crías de ratón, el porteo indujo respuestas calmantes similares a aquellas en los bebéss humanos, a pesar de que los métodos de porteo materno diferían.
Mecanismos sensoriales que provocan las respuestas calmantes
A continuación examinamos las entradas sensoriales requeridas para inducir la respuesta calmante en el modelo de los ratones. En los humanos, se ha sugerido que el tacto materno (15) y el acto de mecer (estimulación vestibular-proprioceptiva) (16) poseen efectos calmantes en los bebés. El hecho de que el agarre del examinador puede provocar la respuesta calmante en los ratones sugiere que en las crías de ratón, la sensación táctil de la piel y la impresión de “estar suspendido” (transporte pasivo del cuerpo) son importantes también. Por otra parte, no parecía que se requirieran inputs olfativos, auditivos y visuales para obtener las respuestas calmantes al porteo manual de crías de ratón (debe indicarse que los párpados de las crías de ratón menores de PND 14 no están abiertos del todo). Para probar la hipótesis en construcción de que el tacto como el materno y la estimulación vestibular-proprioceptiva son importantes también para la respuesta calmante del ratón, primero investigamos el papel de la sensación táctil y encontramos que la respuesta de inmovilización de las crías al porteo manual se veía significativamente reducida por el uso de anestesia local en la piel del cuello dependiendo de la dosis PND 10 (F…) y PND 13 (F…) (Figura 3A). A continuación examinamos el papel de la sensación vestibular y no pudimos encontrar ninguna anormalidad en la respuesta de inmovilización en crías PND 13 que habían recibido daño quirúrgico bilateral al órgano vestibular (laberintectomía) a PND 10 (t…9) (Figura 3B). Después, testamos la privación farmacológica de la propriocepción por sobredosis tóxica de pirodixina (vitamina 6) (17). La inmovilización de las crías de ratón PND 13 por el porteo manual y materno fue atenuada significativamente por la disfunción proprioceptiva (t…) (Figuras 3C y 3D). Estos resultados sugirieron que la sensación táctil y la propriocepción son los inputs sensoriales primarios requeridos para obtener la calma, al menos en los ratones.
Mecanismos neurales de las respuestas cardíacas y motores
A continuación investigamos los mecanismos de salida de cada componente de las respuestas de calma inducidas por el porteo. Consistente con nuestros descubrimientos en losbebés humanos, el índice de variabilidad del ritmo cardíaco rMSSD era considerablemente mayor durante el porteo que durante el sostenido en los ratones (F…) (Figura 4ª y Tabla S3), lo que sugiere la implicación de la activación del nervio parasimpático. Para probar el papel del sistema nervioso autonómico directamente, observamos a las crías de ratón antes y después de la inyección intraperitoneal de receptor muscarinico antagonista atropine, receptor antagonista postináptico B-adrenérgico metoprolol, o solución salina como vehículo. El aumento del intervalo entre latidos provocado por el porteo fue abolido por atropina (sin diferencias entre el porteo y el sostenido, t…, no significativas) (Figuras 4B, 4C y S3), lo que sugiere que el efecto cardíaco del porteo dependía enormemente de la actividad parasimpática. Por otro lado, la respuesta de inmovilidad del porteo era independiente del sistema nervioso autonómico o de la reducción del ritmo cardíaco (Figura 4D). A pesar de que los componentes fisiológicos y del comportamiento en respuesta al porteo son expresados en conjunto, estos componentes pueden ser regulados por circuitos neurales independientes en los animales portados.
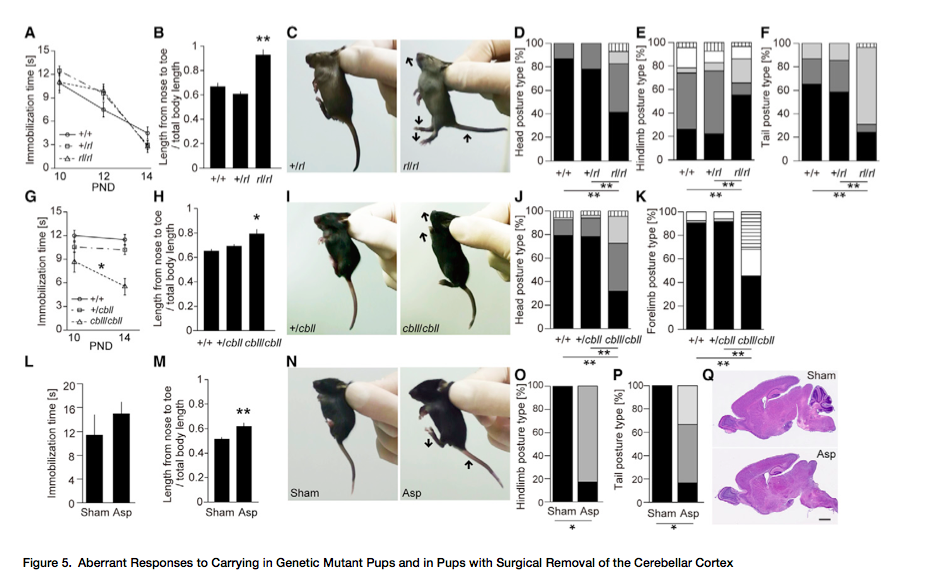
Para explorar los mecanismos subyacentes del cerebro de las respuestas infantiles provocadas por el porteo, examinamos crías de ratón de varias líneas genéticas mutantes que mostraban anormalidades del desarrollo neural e identificamos déficits en dos mutantes, reeler (rl) y cerebelless (dbll). La mutación rl resulta en una supresión parcial de la proteína matriz extracelular Reelin (18) y lleva a hipoplasia severa de la corteza del cerebelo. La mutación cbll causa una expresión reducida de Ptf1a, un factor de transcripción bHLH, que lleva a una pérdida completa de la corteza del cerebelo así como todas las neuronas GABAérgicas en el núcleo del cerebelo (19). Hacia la segunda semana postnatal, la apariencia general y el peso corporal de las crías mutantes homozigeas son indistinguibles de aquellos de tipo salvaje de la misma camada (Figuras S4A y S4B), y la mayoría de las crías sobreviven hasta la maduración sexual a pesar del déficit de coordinación motor en los adultos. La respuesta de inmovilidad al porteo manual fue atenuado progresivamente en las cbll/cbll (F…), pero no en las crías mutantes rl/rl (F…) (Figuras 5A y 5G). En cuanto a regulación postural, tanto las crías rl/rl como las cbll/cbll mostraban anormalidades solapadas durante el porteo manual, incluida la compactación atenuada del cuerpo (reeler, F…) (Figuras 5B y 5H) y dorso-flexión de la cabeza (Test de probabilidad exacto de Fisher) (Figuras 5C, 5D, 5I, y 5J). Además, las crías mutantes rl/rl y cbll/cbll mostraban las posturas de patas traseras (Figura 5E) y cola (Figura 5F) anormales, y posturas de patas delanteras (Figura 5K), respectivamente.
Ya que estos dos mutantes genéticos compartían un fenotipo de malformación congénita de la corteza del cerebelo, examinamos aún más el papel de la corteza del cerebelo en las respuestas provocadas por el porteo en las crías de ratón. Las crías PND 14 que recibieron extirpación quirúrgica de la corteza del cerebelo (Figura 5Q) mostraron regulaciones de postura anormales, parecidas a aquellas de las crías mutantes rl/rl, a saber, el alargamiento del cuerpo (t…) (Figuras 5M y 5N), flexión incompleta de las patas traseras (Test de probabilidad exacto de Fisher) (Figura 5O), y extensión hacia atrás de la cola (Fisher…) (Figura 5P) mientras mostraban una respuesta de inmovilidad intacta (t…) (Figura 5L). Estos descubrimientos sugieren que la característica postura compacta durante el porteo es mediada por la corteza del cerebelo. Por otro lado, la respuesta de inmovilidad puede ser mediada por el olivar inferior y el núcleo pontine, a los que solo afectan los mutantes cbll/cll (19) (Figuras S4C y S4D).
La respuesta calmante facilita el porteo materno
Finalmente, exploramos la importancia funcional de las respuestas identificadas en el porteo materno en las crías de ratón. Propusimos la hipótesis de que si los bebés no cooperaban tranquilizándose y manteniendo una postura compacta en el porteo, la carga materna del porteo aumentaría. Como hemos mostrado en las Figuras 3C y 3D, la disfunción proprioceptiva por el tratamiento con piridoxina inhibió la respuesta de inmovilización durante el porteo. Descubrimos que el tiempo que se requería para el rescate materno (desde el momento en que la madre cogía a la cría hasta el momento en el que la madre y la cría salían de la copa) de las crías tratadas con piridoxina era considerablemente mayor que aquel requerido para las crías tratadas con solución salina (t…) (Figura 3E), lo que sugiere la importancia de la inmovilización para el porteo maternal. Después, para probar el papel de la regulación postural de las crías para el porteo materno, las crías recibieron anestesia general antes de la tarea de rescate materno. Esta intervención dejó a las crías completamente débiles e inmóviles, con todas sus extremidades extendidas (Figura 3F). De nuevo, el tiempo de rescate materno de las crías anestesiadas era considerablemente mayor que aquel de las crías tratadas con solución salina (F…) (Figura 3G). Debe señalarse que las madres rescataron a todas las crías independientemente del tratamiento, lo que indicaba que la motivación maternal de rescatar a las crías no se veía obstaculizada por el tratamiento. Estos datos indicaron colectivamente que ambos aspectos de las respuestas de inmovilización provocadas por el porteo y la regulación postural contribuyeron a facilitar el porteo materno.
Discusión
En una variedad de especies mamíferas como los gatos, leones y ratas, se ha reconocido que el transporte materno oral provoca una postura pasiva y compacta con las patas traseras dobladas en los infantes portados (20-22). Esta regulación postural ha sido estudiada experimentalmente en ratas de laboratorio como “respuesta de transporte” (10,11). Sin embargo, no se han llevado a cabo medidas cuantitativas para la inmovilización durante el porteo, investigación de los aspectos fisiológicos del fenómeno, o la búsqueda del carácter comparativo de este fenómeno en las especies mamíferas. Este estudio es el primero en establecer las sorprendentes similitudes del estado de calma provocado por el porteo entre los bebés humanos y las crías de ratón como una orquestación de movilidad reducida, vocalizaciones angustiadas, y ritmo cardíaco.
En el modelo del ratón, tanto la sensación táctil de la piel en el agarre materno como la propriocepción se requerían para obtener las respuestas calmantes provocadas por el porteo. Este descubrimiento es consistente con literatura anterior que informaba de que el tacto materno y el balanceo rítmico (estimulación vestibular-proprioceptiva) es calmante para los infantes humanos. (15, 16). Sin embargo, el efecto de mecer en las calmantes fisiológicas y en el comportamiento era variable entre los estudios (23, 24). El caminar materno puede ser la estimulación más etimológicamente relevante y proporciona a los infantes con inputs sensoriales calmantes de un modo sinérgico, que podrían ser más efectivos calmando a los bebés que otros tipos de movimiento rítmico tales como mecer de modo mecánico.
Es más, la inmovilización y la adopción de una postura compacta facilitan el porteo materno, como se muestra en las Figuras 3E y 3G. Por lo tanto, las respuestas calmantes pueden aumentar la probabilidad de supervivencia del bebé en casos de escape de emergencia del vínculo madre-bebé y finalmente funcionan apoyando la relación madre-bebé. La conservación de la respuesta calmante en especies mamíferas nidícolas apoya el valor adaptivo de este comportamiento en la relación madre-bebé y, como consecuencia, la supervivencia infantil (10, 20, 22). Curiosamente, Vrugt y Pederson descubrieron que la efectividad de mecer para calmar a los bebés era mayor en la más alta frecuencia de mecer probada (1.5 Hz) (16), lo que sugería aún más la importancia de la respuesta calmante especialmente durante el caminar materno rápido en una emergencia. Sin embargo, en el diseño experimental presente, no controlamos ni medimos la velocidad real del caminar materno. Estudios futuros con medidas precisas de la velocidad del caminar materno se requerirán para caracterizar con precisión sus efectos en calmar a los bebés. Esta respuesta calmante al transporte materno puede desarrollarse incluso antes del nacimiento en los humanos; entre las 36-40 semanas de gestación, los fetos son más activos cuando la madre no es activa en un día (25). Junto con descubrimientos actuales y estos resultados previos, proponemos que la calma provocada por el porteo representa un grupo canónico de respuestas fisiológicas y de comportamiento en las crías de mamíferos nidícolas y funciona en la facilitación de una relación madre-cría eficiente.
El presente estudio proporciona implicaciones inmediatas para las prácticas generales de paternidad. Los efectos identificados del porteo en la activación parasimpática y la reducción del llanto fueron significativos y robustos, para que un breve periodo de porteo pudiera ser un enfoque efectivo para mitigar el llanto causado por irritaciones transitorias tales como las vacunas o ruidos aterradores. Sin embargo, ya que el efecto calmante estaba limitado al periodo real del caminar materno, el infante podía retomar el llanto si la causa subyacente se mantenía después del final del porteo, como el hambre o el dolor crónico. Un entendimiento científico de esta respuesta fisiológica del infante podría prevenir que los padres reaccionaran de forma exagerada al llanto infantil. Este entendimiento sería beneficioso para los padres en reducir la frustración, porque el llanto no mitigado es un factor de riesgo muy importante en el abuso infantil. (26). Además, nuestro simple ensayo de porteo de bebés humanos podría ser utilizado en la evaluación de funciones autonómicas e integraciones sensoriales de desórdenes neurológicos en la infancia temprana, tales desórdenes en el espectro del autismo (ASDs): según los informes, los infantes con ASDs tienen dificultades en el ajuste cooperativo de sus cuerpos al sostenimiento paterno (27). Es más, anormalidades de la estructura del cerebelo (28) y en la integración sensorial (29) están entre los descubrimientos neuropatológicos más consistentes en ASDs. Este estudio también proporciona la implicación para el campo de investigación de la neurociencia. Más obviamente, en cualquier estudio subsecuente que incluya manejo experimental de crías roedoras pre-destete, las respuestas fisiológicas provocadas por el porteo deberían ser reconocidas para evitar influencias autonómicas inesperadas en las medidas subsecuentes que son influenciadas por el sistema nervioso autonómico, como el ritmo cardíaco. Y, de un modo más importante, estos descubrimientos pueden contribuir a la elucidación de los mecanismos neurobiológicos que gobiernan los lazos sociales y la cooperación entre los mamíferos. Anticipamos que el modelo identificado del comportamiento del ratón será una herramienta poderosa para la investigación de mecanismos neurales subyacentes de contribución del bebé al porteo materno y su disfunción en la infancia temprana.
Vídeo sobre el estudio
Traducción de Rebeca Palou del documento original Infant Calming Responses en exclusiva para Mimos y Teta. Prohibida su copia y distribución sin autorización expresa de Nohemí Hervada o Rebeca Palou




Trackbacks/Pingbacks